Строение, свойства и функции нуклеиновых кислот
Содержание:
Строение и функции ДНК
ДНК — полимер, мономерами которой являются дезоксирибонуклеотиды. Модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. Дж. Уотсоном и Ф. Криком (для построения этой модели они использовали работы М. Уилкинса, Р. Франклин, Э. Чаргаффа).
Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т.е. представляет собой двойную спираль (исключение — некоторые ДНК-содержащие вирусы имеют одноцепочечную ДНК). Диаметр двойной спирали ДНК — 2 нм, расстояние между соседними нуклеотидами — 0,34 нм, на один оборот спирали приходится 10 пар нуклеотидов. Длина молекулы может достигать нескольких сантиметров. Молекулярный вес — десятки и сотни миллионов. Суммарная длина ДНК ядра клетки человека — около 2 м. В эукариотических клетках ДНК образует комплексы с белками и имеет специфическую пространственную конформацию.
Мономер ДНК — нуклеотид (дезоксирибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов. Пиримидиновые основания ДНК (имеют в составе своей молекулы одно кольцо) — тимин, цитозин. Пуриновые основания (имеют два кольца) — аденин и гуанин.
Моносахарид нуклеотида ДНК представлен дезоксирибозой.
Название нуклеотида является производным от названия соответствующего основания. Нуклеотиды и азотистые основания обозначаются заглавными буквами.
| Азотистое основание | Название нуклеотида | Обозначение |
|---|---|---|
| Аденин | Адениловый | А (A) |
| Гуанин | Гуаниловый | Г (G) |
| Тимин | Тимидиловый | Т (T) |
| Цитозин | Цитидиловый | Ц (C) |
Полинуклеотидная цепь образуется в результате реакций конденсации нуклеотидов. При этом между 3′-углеродом остатка дезоксирибозы одного нуклеотида и остатком фосфорной кислоты другого возникает фосфоэфирная связь (относится к категории прочных ковалентных связей). Один конец полинуклеотидной цепи заканчивается 5′-углеродом (его называют 5′-концом), другой — 3′-углеродом (3′-концом).
Против одной цепи нуклеотидов располагается вторая цепь. Расположение нуклеотидов в этих двух цепях не случайное, а строго определенное: против аденина одной цепи в другой цепи всегда располагается тимин, а против гуанина — всегда цитозин, между аденином и тимином возникают две водородные связи, между гуанином и цитозином — три водородные связи. Закономерность, согласно которой нуклеотиды разных цепей ДНК строго упорядоченно располагаются (аденин — тимин, гуанин — цитозин) и избирательно соединяются друг с другом, называется принципом комплементарности. Следует отметить, что Дж. Уотсон и Ф. Крик пришли к пониманию принципа комплементарности после ознакомления с работами Э. Чаргаффа. Э. Чаргафф, изучив огромное количество образцов тканей и органов различных организмов, установил, что в любом фрагменте ДНК содержание остатков гуанина всегда точно соответствует содержанию цитозина, а аденина — тимину («правило Чаргаффа»), но объяснить этот факт он не смог.
Из принципа комплементарности следует, что последовательность нуклеотидов одной цепи определяет последовательность нуклеотидов другой.
Цепи ДНК антипараллельны (разнонаправлены), т.е. нуклеотиды разных цепей располагаются в противоположных направлениях, и, следовательно, напротив 3′-конца одной цепи находится 5′-конец другой. Молекулу ДНК иногда сравнивают с винтовой лестницей. «Перила» этой лестницы — сахарофосфатный остов (чередующиеся остатки дезоксирибозы и фосфорной кислоты); «ступени» — комплементарные азотистые основания.
Функция ДНК — хранение и передача наследственной информации.
Строение и функции РНК
РНК — полимер, мономерами которой являются рибонуклеотиды. В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК). Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК.
Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов.
Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой.
Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.
Транспортные РНК содержат обычно 76 (от 75 до 95) нуклеотидов; молекулярная масса — 25 000–30 000. На долю тРНК приходится около 10% от общего содержания РНК в клетке. Функции тРНК: 1) транспорт аминокислот к месту синтеза белка, к рибосомам, 2) трансляционный посредник. В клетке встречается около 40 видов тРНК, каждый из них имеет характерную только для него последовательность нуклеотидов. Однако у всех тРНК имеется несколько внутримолекулярных комплементарных участков, из-за которых тРНК приобретают конформацию, напоминающую по форме лист клевера. У любой тРНК есть петля для контакта с рибосомой (1), антикодоновая петля (2), петля для контакта с ферментом (3), акцепторный стебель (4), антикодон (5). Аминокислота присоединяется к 3′-концу акцепторного стебля. Антикодон — три нуклеотида, «опознающие» кодон иРНК. Следует подчеркнуть, что конкретная тРНК может транспортировать строго определенную аминокислоту, соответствующую ее антикодону. Специфичность соединения аминокислоты и тРНК достигается благодаря свойствам фермента аминоацил-тРНК-синтетаза.
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК: 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК: 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы.
Синтез РНК
Все виды РНК синтезируются при помощи фермента РНК-полимеразы. Она может быть ДНК- и РНК-зависимой, то есть катализировать синтез как на ДНК, так и на РНК-матрице.
Синтез основан на комплементарности оснований и антипараллельности направления чтения генетического кода и протекает в несколько этапов.
Сначала происходит узнавание и связывание РНК-полимеразы с особой последовательностью нуклеотидов на ДНК – промотором, после чего двойная спираль ДНК раскручивается на небольшом участке и начинается сборка молекулы РНК над одной из цепочек, называемой матричной (другая цепочка ДНК называется кодирующей – именно ее копией является синтезируемая РНК). Асимметричность промотора определяет, какая из цепочек ДНК будет служить матрицей, и тем самым позволяет РНК-полимеразе инициировать синтез в правильном направлении.
Следующий этап называется элонгацией. Транскрипционный комплекс, включающий РНК-полимеразу и расплетенный участок с гибридом ДНК-РНК, начинает движение. По мере этого перемещения наращиваемая цепочка РНК постепенно отделяется, а двойная спираль ДНК расплетается перед комплексом и восстанавливается за ним.
Завершающий этап синтеза наступает, когда РНК-полимераза достигает особого участка матрицы, называемого терминатором. Терминация (окончание) процесса может достигаться различными способами.
Химические свойства ДНК.
В воде ДНК образует вязкие растворы, при нагревании таких растворов до 60° С или при действии щелочей двойная спираль распадается на две составляющие цепи, которые вновь могут объединиться, если вернуться к исходным условиям. В слабокислых условиях происходит гидролиз, в результате частично расщепляются фрагменты –Р-О-СН2— с образованием фрагментов –Р-ОН и НО-СН2 , соответственно результате образуются мономерные, димерные (сдвоенные) или тримерные (утроенные) кислоты, представляющие собой звенья, из которых была собрана цепь ДНК (рис. 9).
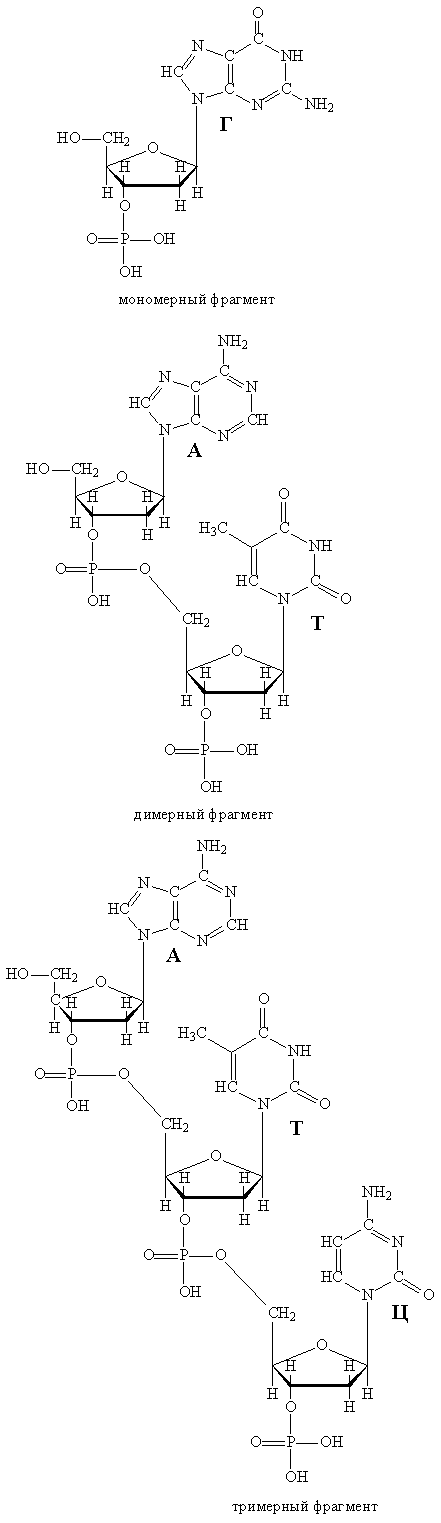
Рис. 9. ФРАГМЕНТЫ, ПОЛУЧАЕМЫЕ ПРИ РАСЩЕПЛЕНИИ ДНК.
Более глубокий гидролиз позволяет отделить участки дезоксирибозы от фосфорной кислоты, а также группировку Г от дезоксирибозы, т.е., более детально разобрать молекулу ДНК на составляющие компоненты. При действии сильных кислот (помимо распада фрагментов –Р(О)-О-СН2-) отщепляются и группировки А и Г. Действие иных реагентов (например, гидразина) позволяет отделить группировки Т и Ц. Более деликатное расщепление ДНК на компоненты проводят с помощью биологического препарата – дезоксирибонуклеазы, выделяемой из поджелудочной железы (окончание —аза всегда указывает на то, что данное вещество представляет собой катализатор биологического происхождения – фермент). Начальная часть названия – дезоксирибонуклеаза – указывает, какое именно соединение расщепляет этот фермент. Все указанные способы расщепления ДНК ориентированы, в первую очередь, на детальный анализ ее состава.
Самая важная информация, содержащаяся в молекуле ДНК, – порядок чередования групп А, Т, Г и Ц , ее получают с помощью специально разработанных методик. Для этого создан широкий набор ферментов, которые находят в молекуле ДНК строго определенную последовательность, например, Ц—T—Г—Ц—A—Г (а также соответствующую ей последовательность на противоположной цепи Г—А—Ц—Г—Т—Ц) и вычленяют ее из состава цепи. Таким свойством обладает фермент Pst I (торговое наименование, оно образуется из названия того микроорганизма Providencia stuartii, из которого получают этот фермент). При использовании другого фермента Pal I удается найти последовательность Г—Г—Ц—Ц. Далее сопоставляются результаты, полученные при действии широкого набора различных ферментов по заранее разработанной схеме, в результате удается определить последовательность таких групп на определенном участке ДНК. Сейчас подобные методики доведены до стадии широкого применения, они используются в самых разнообразных областях, далеких от научных биохимических исследований, например, при идентификации останков живых организмов или установлении степени родства.
Гипотеза РНК-мира
Мир РНК — гипотетический этап эволюционной истории жизни на Земле, в котором самореплицирующиеся молекулы РНК размножались до эволюции ДНК и белков.
Концепция мира РНК была предложена в 1962 году Александром Ричем<, термин был придуман Вальтером Гилбертом в 1986 году. Кроме мира РНК, были предложены и другие химические пути возникновения жизни , и жизнь на основе РНК, возможно, не была первой. Тем не менее, найдено достаточно доказательств возможности существования мира РНК, поэтому гипотеза получила широкое признание.
Подобно ДНК, РНК может хранить и реплицировать генетическую информацию, в виде ферментов — рибозимов она может катализировать (начинать или ускорять) химические реакции, которые имеют решающее значение для существования жизни . Один из наиболее важных компонентов клетки, рибосомы, состоят, в основном, из РНК. Рибонуклеотидные фрагменты во многих коферментах, таких как ацетил-КоА, NADH, FADH и F420, уже давно считаются сохранившимися остатками ковалентно связанных коферментов в мире РНК .
Если РНК-мир существовал, то вероятно, за ним следовала стадия эволюции рибонуклеопротеинов (мир РНП) , которым, в свою очередь, наследовали ДНК и более длинные белки. Причина, по которой ДНК стала преобладающей молекулой хранения генетической информации, может быть связана с тем, что она более стабильна и долговечна, чем РНК . Ферменты-белки, возможно, пришли на смену рибозимам на основе РНК в качестве биокатализаторов, поскольку разнообразие мономеров (аминокислот) делают их более универсальными. Поскольку некоторые кофакторы содержат как нуклеотидные, так и аминокислотные характеристики, может быть, аминокислоты, пептиды и, наконец, белки первоначально были сопутствующими факторами рибозимов.
Нуклеиновые кислоты
Нуклеиновые кислоты — это биополимеры, наряду с белками играющие наиважнейшую роль в клетках живых организмов. Нуклеиновые кислоты отвечают за хранение, передачу и реализацию наследственной информации.
Мономерами нуклеиновых кислот являются нуклеотиды, таким образом они сами представляют полинуклеотиды.
Строение нуклеотидов
Каждый нуклеотид, входящий в состав нуклеиновой кислоты, состоит из трех частей:
- пятиуглеродного сахара (пентозы),
- азотистого основания,
- фосфорной кислоты.
Химические связи между частями нуклеотида ковалентные, образующиеся в результате реакций конденсации (т. е. с выделением молекул воды). Конденсация обратна гидролизу.
В нуклеотиде первый атом углерода пентозы связан с азотистым основанием (связь C-N), а пятый — с фосфорной кислотой (фосфоэфирная связь: C-O-P).
Существуют два основных типа нуклеиновых кислот — ДНК (дезоксирибонуклеиновая кислота) и РНК (рибонуклеиновая кислота). В составе РНК сахар представлен рибозой, а в ДНК — дезоксирибозой. В обоих случаях в нуклеиновых кислотах встречается циклический вариант пентоз. Дезоксирибоза отличается от рибозы отсутствием атома кислорода при втором атоме углерода.
Наличие дополнительной гидроксильной группы (-OH) у рибозы делает РНК молекулой, легче вступающей в химические реакции.
В составе нуклеотидов нуклеиновых кислот обычно встречаются следующие азотистые основания: аденин (А), гуанин (Г, G), цитозин (Ц, C), тимин (Т), урацил (У, U).
Аденин и гуанин относятся к пуринам, остальные — к пиримидинам. В молекуле пуринов имеется два кольца, а у пиримидинов только одно. Урацил почти не встречается в ДНК, а тимин весьма редок для РНК.
То есть для ДНК характерны аденин, гуанин, тимин и цитозин. Для РНК — аденин, гуанин, урацил и цитозин.
Тимин схож с урацилом, отличатся от него лишь метилированным (имеющим группу -CH3) пятым атомом кольца.
Химическое соединение сахара с азотистым основанием называется нуклеозидом. Ниже представлены нуклеозиды, где в качестве сахара выступает рибоза.
Нуклеозид, реагируя с фосфорной кислотой, образует нуклеотид. Ниже представлен нуклеотид, где в качестве сахара выступает дезоксирибоза, а в качестве азотистого основания — аденин.
Именно наличие остатков фосфорной кислоты в молекулах нуклеиновых кислот определяет их кислотные свойства.
Строение нуклеиновых кислот
Нуклеотиды линейно соединяются между собой, образуя длинные молекулы нуклеиновых кислот. Цепочки многих молекул ДНК являются самыми длинными существующими полимерами. Длина молекул РНК обычно существенно меньше ДНК, но при этом различна, т. к. зависит от типа РНК.
При образовании полинуклеотида (нуклеиновой кислоты) остаток фосфорной кислоты предыдущего нуклеотида соединяется с 3-м атомом углерода пентозы следующего нуклеотида. Связь образуется такая же как между 5-м атомом углерода сахара и фосфорной кислотой в самом нуклеотиде – ковалентная фосфоэфирная.
Таким образом, остов молекул нуклеиновых кислот составляют пентозы, между которыми образуются фосфодиэфирные мостики (по-сути остатки пентоз и фосфорных кислот чередуются). От остова в сторону отходят азотистые основания. На рисунке ниже представлена часть молекулы рибонуклеиновой кислоты.
Следует отметить, что молекулы ДНК обычно не только длиннее РНК, но и состоят из двух цепей, соединенных между собой водородными связями, возникающими между азотистыми основаниями. Причем данные связи образуются согласно принципу комплементарности, по которому аденин комплементарен тимину, а гуанин — цитозину.
Подобные связи могут возникать и в РНК (но здесь аденин комплементарен урацилу). Однако в РНК водородные связи образуются между нуклеотидами одной цепи, в результате чего молекула нуклеиновой кислоты сворачивается различным образом.
Состав полимерной цепи нуклеиновых кислот.
Полимерная цепь нуклеиновых кислот собрана из фрагментов фосфорной кислоты Н3РО3 и фрагментов гетероциклических молекул, представляющих собой производные фурана. Есть лишь два вида нуклеиновых кислот, каждая построена на основе одного из двух типов таких гетероциклов – рибозы или дезоксирибозы (рис. 1).
Рис. 1. СТРОЕНИЕ РИБОЗЫ И ДЕЗОКСИРИБОЗЫ.
Название рибоза (от лат. Rib – ребро, скрепка) имеет окончание – оза, что указывает на принадлежность к классу сахаров (например, глюкоза, фруктоза). У второго соединения нет группы ОН (окси-группа), которая в рибозе отмечена красным цветом. В связи с этим втрое соединение называют дезоксирибозой, т.е., рибоза, лишенная окси-группы.
Полимерная цепь, построенная из фрагментов рибозы и фосфорной кислоты, представляет собой основу одной из нуклеиновых кислот –рибонуклеиновой кислоты (РНК). Термин «кислота» в названии этого соединения употреблен потому, что одна из кислотных групп ОН фосфорной кислоты остается незамещенной, что придает всему соединению слабокислый характер. Если вместо рибозы в образовании полимерной цепи участвует дезоксирибоза, то образуется дезоксирибонуклеиновая кислота, для которой повсеместно принято широко известное сокращение ДНК.
История открытия
Фактически открытие дезоксирибонуклеиновой кислоты произошло дважды. Первым открытие молекулы совершил Иоганн Фридрих Мишер в 1869 году. Будучи швейцарским биологом и физиологом, он из клеток, содержащихся в гное, смог выделить большую молекулу с высоким содержанием азота и фосфора. Свое открытие он назвал нуклеин, а позже нуклеиновой кислотой, когда были открыты её кислотные свойства.
Первоначально ученые считали, что основная функция нуклеиновой кислоты в хранении фосфора. А предположения, что она может содержать в себе наследственную информацию, вызывали насмешки, поскольку структура молекулы казалась им слишком простой и однообразной для таких функций. Так же считалось, что наличие дезоксирибонуклеиновой кислоты свойственно только животным клеткам, а в растениях содержится только РНК. Но, в 1934-1935 годах советские ученые-биологи А.Н. Белозерский и А.Р. Кезеля — это наглядно опровергли и опубликовали результаты своих работ в советских и мировых научных журналах.
Повторное открытие ДНК уже в качестве носителя наследственной информации и не только, было совершено в 1944 году. Группа исследователей, состоящая из Освальда Эвери, Колина Маклауда и Маклина Маккарти проводили эксперименты с трансформацией бактерий и доказали, что основную роль в этом процессе играет дезоксирибонуклеиновая кислота .
Дезоксирибонуклеиновая кислота
ДНК это биополимер. В основе мономера ДНК – пентоза. Углевод ДНК является исключением из правил, ведь его формула (C5H10O4) отличается от «нормального» углевода тем, что в ней отсутствует один атом кислорода, поэтому этот углевод получил название «дезоксирибоза».
К остатку дезоксиробозы присоединено одно азотистое основание (цитозин, тимин, аденин и гуанин). Полимерная цепь ДНК образуется путем связывания между собой мономеров. Сшиваются между собой соседние «звенья» остатками фосфорной кислоты, образуя фосфодиэфирную 3’-5’ – связь.
ДНК – это двойная антипараллельная правозакрученная спираль. Две цепи соединены водородными связями, которые возникающими между гетероциклическими соединениями. В ДНК комплементарные пары: A-G и C-T.
Уникальность ДНК в том, что она способна создавать дочернюю молекулу (репликация). Для этого спираль ДНК расходится на две материнские цепи и с помощью ферментов (основной фермент это ДНК-полимераза) на них выстраиваются дочерние цепи, основываясь на правиле комплементарности. В итоге образуется две идентичные друг другу цепи ДНК. Этот процесс обеспечивает безошибочную передачу наследственной информации из поколения в поколение.
Структура РНК
во многом напоминает ДНК, отличие в том, что в основной цепи фрагменты фосфорной кислоты чередуются с рибозой, а не с дезоксирибозой (рис.). Второе отличие – к боковому обрамлению присоединяется гетероцикл урацил (У) вместо тимина (Т), остальные гетероциклы А, Г и Ц те же, что у ДНК. Урацил отличается от тимина отсутствием метильной группы, присоединенной к циклу, на рис. 10 эта метильная группа выделена красным цветом.
Рис. 10. ОТЛИЧИЕ ТИМИНА ОТ УРАЦИЛА – отсутствие у второго соединения метильной группы, выделенной в тимине красным цветом.
Фрагмент молекулы РНК показан на рис. 11, порядок следования группировок А, У, Г и Ц, а также их количественное соотношение может быть различным.
Рис.11. ФРАГМЕНТ МОЛЕКУЛЫ РНК. Основное отличие от ДНК – наличие группировок ОН в рибозе (красный цвет) и фрагмента урацила (синий цвет).
Полимерная цепь РНК приблизительно в десять раз короче, чем у ДНК. Дополнительное отличие в том, что молекулы РНК не объединяются в двойные спирали, состоящие из двух молекул, а обычно существуют в виде одиночной молекулы, которая на некоторых участках может образовывать сама с собой двухцепные спиральные фрагменты, чередующиеся с линейными участками. На спиральных участках взаимодействие пар соблюдается также строго, как в ДНК. Пары, связанные водородными связями и формирующие спираль (А—У и Г—Ц), возникают на тех участках, где расположение групп оказывается благоприятным для такого взаимодействия (рис. 12).
Для подавляющего большинства живых организмов количественное содержание пар А—У больше чем Г—Ц, у млекопитающих в 1,5–1,6 раза, у растений – в 1,2 раза. Существует несколько типов РНК, роли, которых в живом организме различны.
Химическое строение и роль АТФ в энергетическом обмене
Аденозинтрифосфорная кислота (АТФ) содержится в каждой клетке – в гиалоплазме (растворимой фракции цитоплазмы), митохондриях, хлоропластах и ядре.
Она обеспечивает энергией большинство реакций, происходящих в клетке. С помощью АТФ клетка способна двигаться, синтезировать новые молекулы белков, жиров и углеводов, избавляться от продуктов распада, осуществлять активный транспорт и т.п.
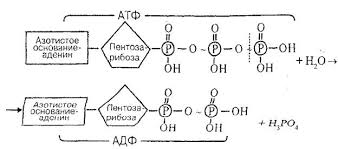
Молекула АТФ образована азотистым основанием, пятиуглеродным сахаром рибозой и тремя остатками фосфорной кислоты. Фосфатные группы в молекуле АТФ между собой соединены высокоэнергетическими (макроэргическими) связями.
В результате гидролитического отщепления конечной фосфатной группы образуется аденозиндифосфорная кислота (АДФ) и освобождается энергия.
После отщепления второй фосфатной группы образуется аденозинмонофосфорная кислота (АМФ) и высвобождается ещё одна порция энергии.
АТФ образуется из АДФ и неорганического фосфата за счёт энергии, которая освобождается во время окисления органических веществ и в процессе фотосинтеза. Называется этот процесс называется фосфориллированием. При этом должно быть использовано не менее 40 кДж/моль АТФ, аккумулированной в её макроэргических связях.
Значит, основное значение процессов дыхания и фотосинтеза состоит в том, что они поставляют энергию для синтеза АТФ, при участии которой в клетке происходит значительное количество различных процессов.
АТФ чрезвычайно быстро восстанавливается.
Пример
У человека каждая молекула АТФ расщепляется и снова возобновляется 2400 раз на сутки, потому средняя длительность её жизни менее 1 мин.
Синтез АТФ осуществляется главным образом в митохондриях и хлоропластах. АТФ, которая образовалась, по каналах эндоплазматического ретикуллюма поступает в те участки клетки, где необходима энергия.
Замечание 8
АТФ играет главную роль в энергетическом обмене клетки и организма.
Любые виды клеточной активности происходят за счёт энергии, которая освобождается во время гидролиза АТФ. Оставшаяся энергия (около 50%), которая освобождается во время расщепления молекул белков, жиров, углеводов и других органических соединений, рассеивается в виде тепла рассеивается и практически существенного значения для жизнедеятельности клетки не имеет.
Функция нуклеиновых кислот
Информация о магазине нуклеиновых кислот, как компьютерный код
Безусловно, наиболее важной функцией нуклеиновых кислот для живых организмов является их роль носителя информации. Поскольку нуклеиновые кислоты могут быть созданы с четырьмя «основаниями» и поскольку «правила спаривания оснований» позволяют «копировать» информацию, используя одну цепь нуклеиновых кислот в качестве шаблона для создания другой, эти молекулы способны как содержать, так и копировать информацию
Поскольку нуклеиновые кислоты могут быть созданы с четырьмя «основаниями» и поскольку «правила спаривания оснований» позволяют «копировать» информацию, используя одну цепь нуклеиновых кислот в качестве шаблона для создания другой, эти молекулы способны как содержать, так и копировать информацию.
Чтобы понять этот процесс, может быть полезно сравнить код ДНК с двоичным кодом, используемым компьютерами. Два кода очень разные по своей специфике, но принцип один и тот же. Так же, как ваш компьютер может создавать целые виртуальные реальности, просто считывая строки 1 и 0, клетки могут создавать целые живые организмы, считывая строки из четырех пар оснований ДНК.
Как вы можете себе представить, без бинарного кода у вас не было бы компьютера и компьютерных программ. Точно так же живые организмы нуждаются в неповрежденных копиях своего «исходного кода» ДНК, чтобы функционировать.
Параллели между генетический код и двоичный код даже побудил некоторых ученых предложить создание «генетических компьютеров», которые могли бы хранить информацию гораздо более эффективно, чем жесткие диски на основе кремния. Однако, поскольку наша способность записывать информацию о кремнии возросла, мало внимания уделялось исследованиям «генетических компьютеров».
Защита информации
Поскольку исходный код ДНК так же важен для клетка поскольку ваша операционная система находится на вашем компьютере, ДНК должна быть защищена от возможного повреждения. Чтобы транспортировать инструкции ДНК в другие части клетки, копии ее информации делаются с использованием другого типа нуклеиновой кислоты – РНК.
Это РНК-копии генетической информации, которые отправляются из ядра и вокруг клетки для использования в качестве инструкций клеточным механизмом.
Клетки также используют нуклеиновые кислоты для других целей. Рибосомы – клеточные машины, которые производят белок – и некоторые ферменты сделаны из РНК.
ДНК использует РНК как своего рода защитный механизм, отделяющий ДНК от хаотической среды цитоплазма, Внутри ядра ДНК защищена. За пределами ядра движения органелл, везикул и других клеточных компонентов могут легко повредить длинные и сложные нити ДНК.
Тот факт, что РНК может действовать как в качестве наследственного материала, так и в качестве фермента, подтверждает идею о том, что самой первой жизнью могла быть самореплицирующаяся, самокатализирующаяся молекула РНК.
ДНК
Дезоксирибонуклеиновая кислота является макромолекулой. С ее помощью происходит процесс передачи и реализации генетической информации. Эти сведения необходимы для программы развития и функционирования живого организма. У животных, растений, грибов ДНК входит в состав хромосом, находящихся в ядре клетки, а также находится в митохондриях и пластидах. У бактерий и архей молекула дезоксирибонуклеиновой кислоты цепляется за клеточную мембрану с внутренней стороны. В таких организмах присутствуют в основном кольцевые молекулы ДНК. Они получили название «плазмиды». По химическому строению дезоксирибонуклеиновая кислота представляет собой полимерную молекулу, состоящую из нуклеотидов. Эти компоненты, в свою очередь, имеют в своем составе азотистое основание, сахар и фосфатную группу. Именно за счет двух последних элементов образуется связь между нуклеотидами, создавая цепи. В основном макромолекула ДНК представлена в виде спирали из двух цепей.
Транскрипция
Синтез всех молекул происходит во время транскрипции, то есть переписывании генетической информации с определенного оперона ДНК. Процесс в некоторых моментах похож на репликацию, а в других существенно отличается от нее.
Сходствами являются следующие части:
- начало идет с деспирализации ДНК;
- происходит разрыв водородных связей между основаниями цепей;
- к ним комплементарно подстраиваются НТФ;
- происходит образование водородных связей.
Отличия от репликации:
- при транскрипции расплетается лишь участок ДНК, соответствующий транскриптону, в то время как при репликации расплетению подвергается вся молекула;
- при транскрипции подстраивающиеся НТФ содержат рибозу, и вместо тимина урацил;
- информация списывается лишь с определенного участка;
- после образования молекулы водородные связи и синтезированная цепь разрываются, а цепь соскальзывает с ДНК.
Для нормального функционирования первичная структура РНК должна состоять только из списанных с экзонов ДНК-участков.
У только что образованных РНК начинается процесс созревания. Молчащие участки вырезаются, а информативные сшиваются, образуя полинуклеотидную цепь. Далее, каждый вид имеет присущие только ему превращения.
В и-РНК происходит присоединение к начальному концу. К конечному участку присоединяется полиаденилат.
В т-РНК модифицируются основания, образуя минорные виды.
У р-РНК также метилируются отдельные основания.
Защищают от разрушения и улучшают транспортировку в цитоплазму белки. РНК в зрелом состоянии с ними соединяются.